
|

|

|
|
|
|
|
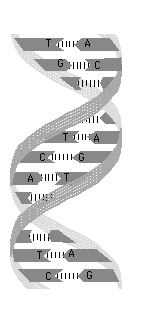 1953年にワトソン(James D. Watson)とクリック(Francis H.C. Crick)は、右図にあるDNAの二重らせん(double helix)構造モデルを提唱しました。このモデルはDNA繊維のX線回折データをもとに、どんなDNAでもアデニン(A)とチミン(T)の含量が等しく、グアニン(G)とシトシン(C)の含量が等しいという実験事実を手がかりとして作られました。DNAは2本の鎖でできていること、1本の鎖はA、T、G、Cの4種類の塩基が並んでできており、その並び方すなわち配列(sequence)に遺伝情報が蓄えられていること、そしてAとT、CとGがそれぞれペアとしてうまくかみあう相補的な(complementary)立体構造をしていることから、鎖の一方の並びが与えられれば他方は自動的に決まることが示されました。ワトソン・クリックの二重らせんモデルは、相補構造によるDNAの複製メカニズムを明らかにし、ここに遺伝という生命の基本問題の分子的理解が可能となったのです。1953年は生命現象を分子レベルで解明しようという分子生物学の始まりでもありました。しかしながら、実際にA、T、G、Cの並びを読み取る技術ができるようになるには、さらに20年以上が必要となります。
1953年にワトソン(James D. Watson)とクリック(Francis H.C. Crick)は、右図にあるDNAの二重らせん(double helix)構造モデルを提唱しました。このモデルはDNA繊維のX線回折データをもとに、どんなDNAでもアデニン(A)とチミン(T)の含量が等しく、グアニン(G)とシトシン(C)の含量が等しいという実験事実を手がかりとして作られました。DNAは2本の鎖でできていること、1本の鎖はA、T、G、Cの4種類の塩基が並んでできており、その並び方すなわち配列(sequence)に遺伝情報が蓄えられていること、そしてAとT、CとGがそれぞれペアとしてうまくかみあう相補的な(complementary)立体構造をしていることから、鎖の一方の並びが与えられれば他方は自動的に決まることが示されました。ワトソン・クリックの二重らせんモデルは、相補構造によるDNAの複製メカニズムを明らかにし、ここに遺伝という生命の基本問題の分子的理解が可能となったのです。1953年は生命現象を分子レベルで解明しようという分子生物学の始まりでもありました。しかしながら、実際にA、T、G、Cの並びを読み取る技術ができるようになるには、さらに20年以上が必要となります。
一方のタンパク質は20種類のアミノ酸が一次元的につながった鎖状の高分子であり、立体的には個々のタンパク質ごとに異なる複雑な構造をしています。アミノ酸の並び、すなわち一次構造は、1953年にサンガー(Frederick Sanger)が初めてインシュリンで決定しました。立体構造については、1960年にケンドルー(John C. Kendrew)らがX線結晶解析によりミオグロビンを解明しました。ミオグロビンはαヘリックス(α-helix)と呼ばれるらせん構造が6本集まってできていますが、αヘリックスについては1951年にポーリング(Linus Pauling)がホモポリアミノ酸(同種類のアミノ酸からなる鎖)でのモデル構造から、タンパク質での存在を予言していたものです。その後、多くのタンパク質の立体構造が解明されるにつれ、タンパク質の機能、例えば酵素(enzyme)が基質(substrate)を認識して作用する機能は、立体構造の相補性に基づいていることが明らかになってきました。立体構造の相補性とは、鍵と鍵穴のような形としての相補性だけでなく、DNA二重らせんを安定化させる水素結合のように、化学的性質の相補性が重要です。DNAもタンパク質も一次元的な鎖から形成される生体高分子ですが、生体内では三次元的にきまった形をとって初めて生物的な働きをします。それぞれの分子の特異的な立体構造が、特異的な相互作用につながっているのです。